Computational, Systems and Developmental Neuroscience
Axon guidance
During neural development, growing nerve fibres (axons) use molecular cues in their environment in order to navigate to their targets. One of the most important guidance signals they are believed to use are concentration gradients, i.e. chemotaxis. We are studying how axons detect and respond to such gradients, using a combination of mathematical modelling, in vitro and in vivo experiments. Below is a summary of some of our work from the past few years.
- Can gradients wire the brain? Although textbook descriptions of axon guidance invoke concentration gradients, quantitative arguments suggest that such gradients in the brain might actually be too shallow to provide effective guidance. See Goodhill, 2016 in Trends in Neurosciences.
- Fundamental limits to chemosensation. Concentration measurement by any sensing device is constrained by several sources of noise. Using the fluctuation-dissipation theorem we recently derived how these fundamental limits vary across dimensions, and in particular how 2D sensing (e.g. where ligands are bound to a surface) is different from 3D sensing. See Bicknell et al 2015 in Nature Communications. This was a collaboration with Peter Dayan.
- Modelling signal transduction underlying axon growth and guidance. In Bicknell & Goodhill 2018 we proposed a mathematical model that couples growth and guidance at the level of signal transduction, and showed that it can quantitatively explain this dataset of axon growth and guidance under precisely controlled gradient conditions. In Padmanabhan & Goodhill 2018 we showed through modelling how the molecular interaction network involved in axon growth can exhibit bistability, with one state representing a fast-growing growth cone and the other a paused growth cone.
- Modal gating Concentration sensing relies on ion channels. By mathematically modeling the basic biophysical events that control ion channel opening, we introduced a new principle for understanding the origin of modal gating. See Bicknell & Goodhill 2016 in PNAS.
- Optimal receptor placement. Where would be the best place to put receptors on a sensing device to maximize the information available about an external gradient, and how is gradient sensing affected by receptor diffusion? We recently derived answers to these questions using tools from information theory: see Nguyen et al 2014a and Nguyen et al 2014b. This was a collaboration with Peter Dayan.
- Microfluidic gradient generators. Microfluidics is a powerful new class of technologies for precisely manipulating small volumes of fluid. In collaboration with Justin Cooper-White we are developing microfluidics-based devices for examining the response of axons to gradients.
- Calcium signalling in axon guidance. Calcium plays a critical role in the signalling pathways underlying axonal chemotaxis, recently reviewed in Trends in Neurosciences (Sutherland et al 2014). We modelled this mathematically in a paper in Neuron (Forbes et al, 2012), and are now developing this modelling further.
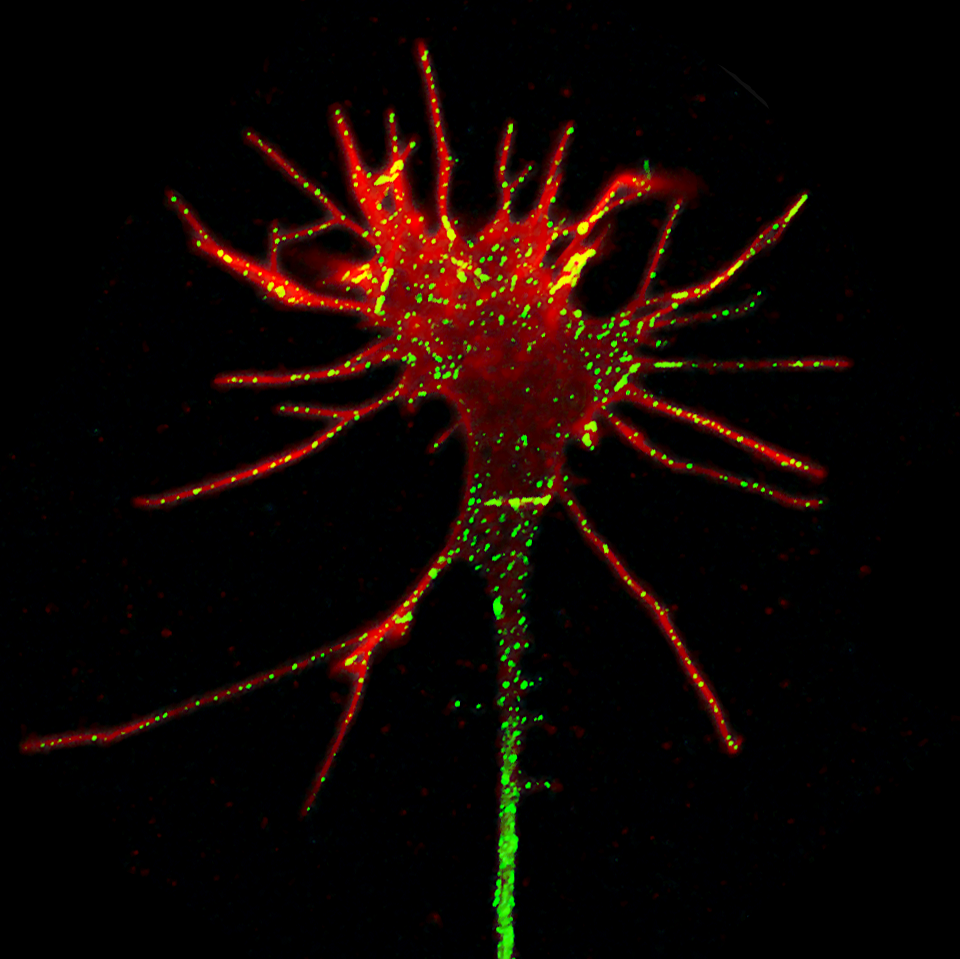
- Growth cone shape analysis Growth cones at the tips of axons have a complex and dynamic morphology. We recently extracted the principal components of growth cone shape variation from a large set of timelapse movies of growth cones, and found that growth cone shape oscillates with an average period of about 30 minutes. A simple mathematical model of dynamic microtubule instability was able to explain this periodicity. See our recent paper in BMC Biology (Goodhill et al, 2015).
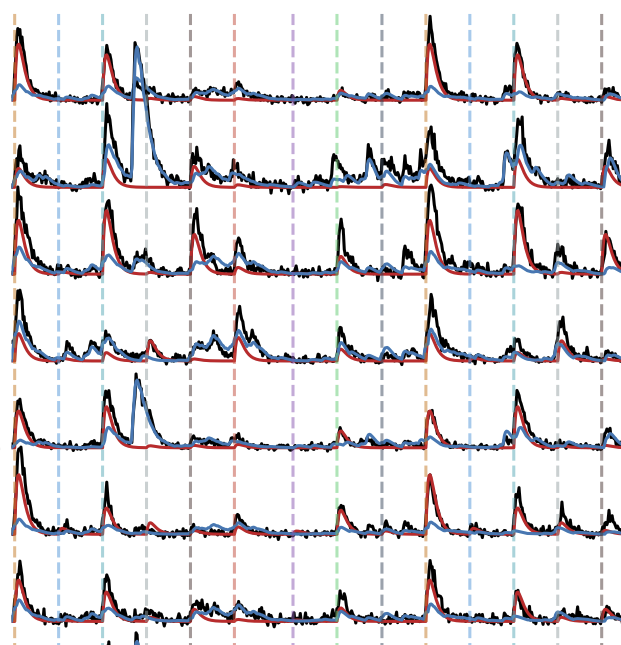
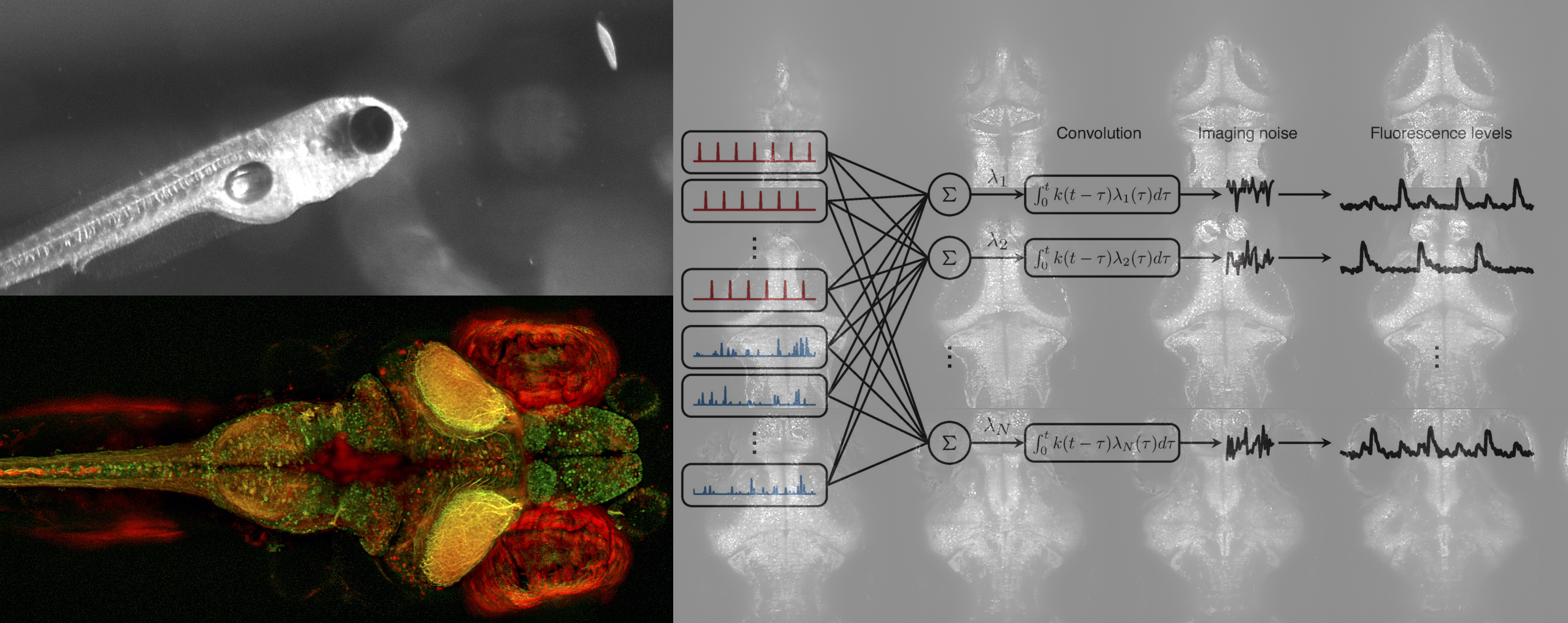
- Axon guidance in zebrafish tectum. Because larval zebrafish are transparent it is possible to visualize retinal axons as they search for targets in the optic tectum. We recently found that axonal branching plays an unexpectedly key role in this process (see Simpson et al, 2013), and this was not strongly affected affected by neural activity (see Kita et al, 2015a). We recently reviewed retinotectal map formation in zebrafish in Kita et al, 2015b. This was a collaboration with Ethan Scott.
- Axon targetting in the corpus callosum. We recently collaborated with Linda Richards on a paper in Neuron (Saurez et al, 2014) which showed that balanced interhemispheric cortical activity is required for correct targeting of axons in the corpus callosum.
Some older papers
- Yuan, J., Chan, S., Mortimer, D., Nguyen, H. & Goodhill, G.J. (2013). Optimality and saturation in axonal chemotaxis. Neural Computation, 25, 833-853 (2013). PDF
- Forbes, E.M., Hunt, J.J. & Goodhill, G.J. (2011). The combinatorics of neurite self-avoidance. Neural Computation, 23, 2746-2769. PDF
- Thompson, A.W., Pujic, Z., Richards, L.J. & Goodhill, G.J. (2011). Cyclic nucleotide-dependent switching of mammalian axon guidance depends on gradient steepness. Molecular and Cellular Neuroscience, 47, 45-52. PDF
- Mortimer, D., Dayan, P., Burrage, K. & Goodhill, G.J. (2011). Bayes-optimal chemotaxis. Neural Computation, 23, 336-373. PDF
- Mortimer D, Pujic Z, Vaughan T, Thompson AW, Feldner J, Vetter I, Pujic Z, & Goodhill GJ (2010). Axon guidance by growth rate modulation. Proc. Natl. Acad. Sci. USA, 107, 5202-5207. PDF SI
- Mortimer, D., Dayan, P., Burrage, K. & Goodhill, G.J. (2010). Optimizing chemotaxis by measuring unbound-bound transitions. Physica D, 239, 477-484. PDF F1000 review
- Mortimer D, Feldner J, Vaughan T, Vetter I, Pujic Z, Rosoff WJ, Burrage K, Dayan P, Richards LJ, Goodhill GJ (2009). A Bayesian model predicts the response of axons to molecular gradients. Proc. Natl. Acad. Sci. USA, 106, 10296-10301. PDF SI
- Mortimer, D., Fothergill, T., Pujic, Z., Richards, L.J. & Goodhill, G.J. (2008). Growth Cone Chemotaxis. Trends in Neurosciences, 31, 90-98. PDF
- Xu, J., Rosoff, W.J., Urbach, J,S. & Goodhill, G.J. (2005). Adaptation is not required to explain the long-term response of axons to molecular gradients. Development, 132, 4545-4552. PDF
- Rosoff, W.J., Urbach, J.S., Esrick, M., McAllister, R.G. Richards, L.J. & Goodhill, G.J. (2004). A new chemotaxis assay shows the extreme sensitivity of axons to molecular gradients. Nature Neuroscience, 7, 678-682. PDF News and Views F1000 reviews